FASE METABÓLICA DE ABSORÇÃO:
O melhor modo de obter um entendimento das inter-relações das vias metabólicas é
conhecer as mudanças no metabolismo durante o ciclo jejum-alimentação. É importante
lembrar que o intestino não é um reservatório que o organismo pode ir retirando nutrientes
quando precisa. As refeições são digeridas numa velocidade que depende da constituição
química da dieta, independente da necessidade nutricional do animal. Então, o organismo
absorve os nutrientes em períodos metabólicos que geralmente não coincidem com o gasto
desses nutrientes. Sendo assim, deve haver um sofisticado sistema para manter a
homeostase dos nutrientes.
Vamos falar dos principais nutrientes supridores de
energia que são a glicose, os aminoácidos, os ácidos graxos e os corpos cetônicos.
Chamamos esses nutrientes supridores de energia de combustíveis metabólicos e os
mecanismos fisiológicos para manutenção do suprimento de combustíveis relacionados
com a sua demanda é conhecido como o homeostasia de combustíveis, que é mantida por
diversos mecanismos incluindo o eixo insulina-glucagon, o eixo hipotalâmico-hipofisário e
no sistema nervoso central. Para estes de combustíveis metabólicos existem várias formas
de armazenamento.
Estes
compostos podem ser direcionados para o ciclo de Krebs para que ocorra então a produção
de energia. Importante lembrar que os aminoácidos também fornecem esqueletos
carbônicos que vão para o CK. Assim todos esses combustíveis metabólicos podem ser
oxidados e convertidos a acetil-coa que depois é queimada no ciclo de Krebs.
Em alguns
esquemas nós vamos ver uma fogueirinha, ela representa a Fornalha que exatamente o
ciclo de krebs (CK). Nesse esquema nós temos as inter-relações metabólicas que ocorrem
durante o estado bem alimentado que também podemos chamar de fase metabólica de
absorção. As três principais vias são glicólise, beta-oxidação e ciclo de Krebs.
No momento metabólico de absorção os eventos metabólicos no fígado e nos órgãos
periféricos direcionam os nutrientes para armazenagem. Os aminoácidos e a
glicose seguem um caminho diferente dos TG. Isso porque os aas e a glicose são
absorvidos pelos capilares que drenam o intestino e vão se reunir para formar então a veia
porta e levar esses nutrientes recém-absorvidos diretamente para o fígado através da veia
porta. Dessa forma a drenagem feita no intestino pela veia porta leva glicose e aas para o
fígado, enquanto que os triglicerídeos são absorvidos na forma de quilomicrons através do
sistema linfático. Eles só vão cair na circulação sistêmica na altura da veia cava superior.

Essa circulação entero-hepática ela é organizada de modo que todos os nutrientes
absorvidos no intestino que vão para o sangue passem primeiro pelo fígado antes de
penetrar na circulação sistêmica (isso coloca o fígado numa posição de sentinela onde ele
pode alterar a constituição do sangue Portal antes do sangue ser distribuído para os outros
tecidos).
Nesse esquema está representada a disposição da glicose, aminoácidos e gordura por
vários tecidos no estado bem alimentado, ou seja, durante a fase metabólica de absorção. A
glicose é captada pelo fígado e uma boa parte dessa glicose que entra no fígado acaba
sendo armazenada como glicogênio. Sendo que o fígado consegue armazenar cerca de
10% do seu peso total como glicogênio. Lembrando que para um fígado humano isso seria
o equivalente a aproximadamente 100 gramas, Entretanto essa capacidade de 100 g não
vai acomodar toda a glicose que é captada pelo fígado durante essa fase de absorção, isso
quando a gente faz uma refeição rica em carboidratos. Então deve haver um mecanismo
adicional para a deposição desse excesso de glicose, e esse mecanismo alternativo é
exatamente a síntese de gordura, a síntese de ácidos graxos.
Essa síntese de ácidos
graxos ocorre porque neste momento metabólico a glicólise está ativa gerando piruvato que
então é convertido em acetil-coa dentro da mitocôndria para que esse acetil-coa possa
entrar no ciclo de Krebs. Entretanto nesse momento metabólico a atividade do ciclo de
Krebs já está mais do que suficiente para suprir as necessidades do fígado. Assim, ocorre
um excesso de acetil-coa que é combinado oxaloacetato para formar citrato e esse citrato é
transportado para o citosol e lá no citosol o citrato vai contribuir com 2 carbonos na forma de
acetil-coa para a síntese de ácidos graxos, sendo que o oxaloacetato depois retorna para a
mitocôndria para formar piruvato.
Esse excesso de carboidratos que é convertido em ácido
graxo é uma reação irreversível. Portanto, o excesso de carboidratos pode virar gordura,
mas a gordura não pode virar carboidrato, a não ser que a gente esteja falando de um ácido
graxo com número ímpar de carbono, daí ele fornece o esqueleto de carbono para o CK pra
ser convertido em glicose na gliconeogenese
Esses ácidos graxos recém sintetizados no
fígado são empacotados como VLDL ou lipoproteínas de muito baixa densidade, para isso
os ácidos graxos são inicialmente esterificados em TG e depois recebem uma cobertura de
fosfolipídeos e colesterol além claro de proteínas e então vão formar as proteínas de muito
baixa densidade que serão exportadas para os tecidos extra-hepáticos. Além de ser
armazenada como glicogênio e o excesso ser convertido em ácidos graxos a glicose pode
ser consumida na via glicolítica ou ainda na Via das Pentoses.
Mas o fígado não capta toda
a glicose que é absorvida. Uma parte da glicose segue pela circulação sistêmica e será
utilizada pelos outros tecidos como o cérebro que é um órgão que quase que
completamente depende de glicose para produção de ATP. Além disso, eritrócitos e medula
renal só podem fazer glicólise. Nesse momento metabólico a glicose que ganha o tecido
adiposo será convertido em glicerol para a formação dos triacilgliceróis que serão
armazenados no tecido adiposo. Os músculos também captam essa glicose e vão
convertê-la em glicogênio ou queimá-la para obtenção de energia dependendo da situação
metabólica do músculo.
Para introduzir a discussão acerca do metabolismo de aminoácidos durante a fase de
absorção podemos classificar os aminoácidos de acordo com as suas características
metabólicas como mostrado nesta tabela que então nós temos dois principais grupos de
aminoácidos que são os aminoácidos essenciais e os aminoácidos não essenciais. Dentro
do grupo dos aminoácidos não essenciais nós temos Glutamato, Aspartato e Alanina que
são importantes aminoácidos de transporte; e dentro do grupo dos aas essenciais teremos
Leucina, Isoleucina e Valina que formam um grupo especial de aas conhecido como Aas de
cadeia ramificada (ou BCAA).

Parte dos aminoácidos que são absorvidos na mucosa
intestinal são utilizados pelos próprios enterócitos para obtenção de energia, mas a maior
parte segue para o sangue portal para então ser distribuído. Também podem acontecer
transformações de aas dentro dos enterócitos durante o processo de absorção. Isso
significa que alguns aminoácidos podem ser convertidos na forma de transporte, e justifica
fato do perfil de aminoácidos na veia porta ser consideravelmente diferente do perfil de
aminoácidos da dieta, sendo que o principal aas no sangue Portal é exatamente a Alanina
que é o principal aas de transporte.
Quando o sangue Portal alcança o fígado uma grande
quantidade de aas é removida do sangue e apenas um pouco mais de 20% desses
aminoácidos que estão chegando ao fígado durante a fase de absorção é que vão passar
para a circulação sistêmica, assim a concentração de aminoácidos no sangue, assim como
também é mantida a concentração de glicose né, é mantida uma concentração
relativamente constante de aas durante a fase de absorção. Podemos observar que o fígado é um importante local de síntese de proteínas, seja
proteínas para repor proteínas do próprio fígado ou ainda proteínas sintetizadas que vão ser
liberadas no plasma que são proteínas séricas. Como podemos observar a maioria dos
aminoácidos absorvidos pelo fígado acaba sendo oxidada

Para isso inicialmente ocorre a
perda do grupamento amino que é tóxico e precisa ser então convertido em ureia no ciclo
da ureia, e o alfa-cetoácido correspondente é direcionado ao CK (percorre as vias
oxidativas semelhante dos carboidratos para que possam ser completamente oxidados e
fornecer energia) ou ainda esses esqueletos carbônicos podem ser convertidos em glicose
na via de gliconeogênese, mas não durante a fase de absorção normalmente a não ser que
tenha sido feita uma dieta restritiva em carboidratos.

Vale lembrar que os aminoácidos eles
podem suprir o ciclo de Krebs com esqueletos carbônicos, diferentemente do acetil-coa que
é incapaz de suprir o CK com esqueleto carbônico porque os dois carbonos que entram do
acetil-coa saem das reações de descarboxilação e essa capacidade de usar aminoácidos
para suprir o CK ou até mesmo produzir glicose é muito importante nos carnívoros
verdadeiros porque nesses animais a dieta natural contém grande quantidade de proteína e
pouco carboidrato. Entretanto a necessidade de glicose nesses animais não é menor do
que nos outros animais, daí a necessidade dessa conversão.
Como falado, um pouco mais de 20% desses aminoácidos escapam captação hepática e
são direcionados para os tecidos extra-hepáticos.
Boa parte desses aminoácidos que
escapam são aminoácidos de cadeia ramificada. Essa rota é geralmente seguida pelos
aminoácidos essenciais. Os aminoácidos então ganham o músculo onde passam a fazer
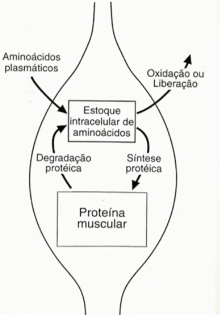
parte de um estoque intracelular de aminoácidos.
Vale lembrar que nessa fase de absorção
tem uma secreção maior de insulina que irá promover tanto a captação de glicose como
também a captação de aminoácidos pelo músculo. Então essa glicose que é captada pelo
músculo é convertida em glicogênio muscular que será utilizado para o trabalho muscular.
Já os aas vão compor esse estoque intracelular de aas que é direcionado para a síntese
proteica sempre que o toque intracelular estiver alto ocorre o favorecimento da síntese
proteica. quando esse estoque for diminuído ocorrerá a degradação proteica, isso não
durante a fase de absorção, mas sim em outra fase. Então que a gente pode considerar é
que essa proteína muscular tem papel funcional não apenas na locomoção e postura, mas
também como um depósito de aas.

Ainda na fase de absorção é importante a gente falar
dos triacilgliceróis que chegam da dieta seja na forma de quilomícrons ou que foram
recém-sintetizados no fígado e empacotados como VLDL. Tanto os quilomicrons como as
lipoproteínas de muito baixa densidade vão transitar pela circulação alcançando tecidos que
se utilizam de ácidos graxos como o tecido muscular, o tecido adiposo e também a glândula
mamária em lactação, e tanto o quilomícron quanto o VLDL, eles possuem uma proteína
chamada C2 que está na superfície do quilomícron e na superfície do VLDL.
Essa proteína
C2 que está presente na superfície da lipoproteína quilomicron e também do VLDL, estimula
então a ação da lipase lipoproteica presente no endotélio dos vasos que banham esses
tecidos adiposo, músculo e glândula mamária em lactação. Então a Lipase Lipoproteica
cliva os TG dessas lipoproteínas liberando ácidos graxos e glicerol que são absorvidos
pelos tecidos e lá resterificados e armazenados.
Essa Lipase Lipoproteica nos vasos que
banham os tecidos adiposo está estimulada pela ação da insulina, então durante a fase de
absorção os ácidos graxos tanto dos triacilgliceróis que estão nos quilomícrons como os
ácidos graxos dos TG das VLDL são seletivamente transferidos para o tecido adiposo.
Recapitulando: durante essa fase de absorção os
aminoácidos e carboidratos que estão em excesso são convertidos em ácidos graxos que
vão transitar na forma de VLDL e esses VLDLs alcançam o tecido adiposo e são
armazenados aí. Além disso, aqueles ácidos graxos que foram absorvidos pelos enterócitos
na forma de quilomícrons também são esvaziados pelo tecido adiposo e como se não fosse
o bastante, a insulina também direciona a captação de glicose pelo tecido adiposo, e essa
glicose é usada para a síntese do glicerol que será utilizado no processo de resterificação
dos TG para armazenamento de gotículas de gordura, ou ainda pode ser utilizado para
síntese de mais ácidos graxos e assim a armazenagem da gordura no tecido adiposo.
FASE DE PÓS-ABSORÇÃO:
Nesse momento de pós-absorção teremos uma diminuição da taxa de absorção de glicose
e isso reflete também na diminuição da concentração plasmática de glicose. Isso vai
remover o estímulo para secreção de insulina (insulina no sangue tbm diminui) e em
contrapartida aumenta a secreção de glucagon que vai interferir com as enzimas hepáticas.
A fase de pós-absorção é um período relativamente breve (em geral poucas horas) e que
ocorre entre as refeições em animais bem alimentados.
Ela se caracteriza por variações de
curto prazo que mobilizam os nutrientes dos locais de depósito para manter a
disponibilidade de combustível para os tecidos metabolicamente ativos. Ou seja, é
caracterizada por essa mobilização dos combustíveis para fora dos locais de
armazenamento para que possam ser utilizados.
Com a diminuição da insulina e aumento
da secreção de glucagon característico da fase de pós-absorção também chamada de jejum
inicial nós temos a ação do próprio hormônio glucagon.
O glucagon interage com o receptor
do hepatócito e estimula através de uma cascata de sinalização celular a fosforilação de
várias enzimas. Então nós temos as 4 enzimas-chaves que estão fosforiladas sobre a
influência do AMPc que é o segundo mensageiro dessa sinalização mediada pelo glucagon.
Logo, essa capacidade de regulação enzimática pela fosforilação é um tipo covalente de
modulação e nós temos que as enzimas glicogênio fosfatase e as enzimas frutose
1-6-bifosfatases estão ativadas durante a fosforilação.
Já a glicogênio sintetase e a
fosfofrutoquinase estão inibidas pela fosforilação. Portanto, esse momento metabólico é um
momento aonde nós temos a degradação do glicogênio, ou seja, a
glicogenólise, e essa
glicose liberada é encaminhada para o sangue no intuito de equilibrar a glicemia e a via de
gliconeogênese também está ativa e também já vai suprir a demanda de glicose que ocorre
principalmente no final dessa fase de pós-absorção.
Além dessa mobilização de
carboidratos no fígado o músculo também vai reagir a essa demanda metabólica de glicose
e nesse caso ele vai reagir mobilizando os aas para garantir essa gliconeogênese hepática.
O que acontece é que com declínio da concentração de insulina no sangue ocorrem em
dois efeitos no músculo: primeiro é que a própria entrada de aminoácidos que vem do
sangue vai ser diminuída; e segundo a entrada de glicose também vai estar diminuída.
Então o que acontece é que como diminui o “pow” intracelular de aas, inicia-se a
degradação das proteínas para não ter o “pow” intracelular de aminoácidos; e como não tá
entrando glicose isso resulta na utilização de aminoácidos para manter a energia do próprio
tecido muscular (esse evento é chamado proteólise muscular que ocorre durante essa fase
de pós-absorção que também é chamada de jejum inicial)
Logo,a
fase de pós-absorção ocorre proteólise muscular que vai tanto suprir as necessidades
energéticas do músculo como também fornecer esqueletos carbônicos para o fígado para
que ele possa utilizar esse esqueleto carbônico na via de gliconeogênese e produzir a
glicose nova que é biodisponibilizada para o organismo.

É importante a gente lembrar que
as fontes primárias de aminoácidos disponibilizados pelos músculos são exatamente
aqueles BCAA ou AACR (que são aas de cadeia ramificada - Leucina, Isoleucina e Valina).
Esses aas são utilizados durante esse evento de proteólise mesmo porque esses
aminoácidos contribuem com aproximadamente um terço de todos os aminoácidos
musculares. Então o catabolismo dos aminoácidos cadeia ramificada vai começar com a
transaminação (o grupo Amina é transferido para um outro alfacetoacido, como o
Oxoglutarato, por exemplo, formando o glutamato; e o alfa-cetoácido doa os carbonos para
o ciclo de Krebs que então fornece piruvato que é aminado formando a Alanina e essa
Alanina que é exportada para o sangue).
O resultado final então é a convenção dos
aminoácidos de cadeia ramificada em Alanina que ganha a circulação e então é captada
pelo fígado e direcionada para a gliconeogênese.
Esse mecanismo pode parecer um pouco
complicado né? Então por que transformar em alanina e não encaminhar o próprio AACR para
circulação?
A gente tem que se lembrar que o fígado não capta esses aminoácidos de
cadeia ramificada, já a Alanina ele capta bem. É importante lembrar que essa mobilização
de aminoácidos pelo músculo é influenciado em grande parte pela falta de insulina mas o
hormônio adrenocortical cortisol também tem grande efeito na estimulação dessa
degradação proteica. Em relação ao metabolismo de ácidos graxos nós temos que durante
a fase de pós-absorção ocorre que aqueles ácidos graxos que estavam armazenados no
tecido adiposo acabam sendo liberados. Esse efeito se dá pela ação da enzima lipase
hormônio sensível também chamada de triacilglicerol lipase. Essa enzima é ativada por uma
proteína kinase-A resultante da estimulação do segundo mensageiro AMPc.
Como o
glucagon parece ter um efeito mais direcionado para o fígado os hormônios que
provavelmente estão estimulando essa sinalização são a adrenalina e a noradrenalina, mas
eu não descarto a sinalização do glucagon também no tecido adiposo. Os ácidos graxos
liberados pela ação da lipase hormônio sensível vão então para circulação onde serão
transportados pela Albumina. Diferente dos triglicerídeos que são transportados pelos
quilomicrons e pelas VLDLs, esses ácidos graxos não estão esterificados eles são ácidos
graxos livres e são então incorporados pela Albumina e transportados até a sua utilização.
Jejum:
Nessa fase metabólica chamada de jejum ocorre diminuição plasmática tanto da
concentração de glicose como da concentração de aminoácidos. Isso reflete na secreção
de insulina que é diminuída e no aumento da secreção de glucagon e essa mobilização
hormonal tem duas grandes prioridades: a primeira é manter os níveis de glicose
plasmática; e a segunda é mobilizar os ácidos graxos do estoque promovendo a lipólise e
assim poder utilizar esses ácidos graxos como energia seja na forma de ácidos graxos ou
ainda como corpos cetônicos.

Como já discutido anteriormente, os aas formam um
importante depósito de precursores da glicose e de substratos precursores de energia.
Entretanto, durante o jejum ou subnutrição prolongada não seria vantajoso para os animais
depender muito de seus músculos esqueléticos como fonte de energia porque isso poderia
culminar em fraqueza intensa por consumo das proteínas musculares. Então existem
mecanismos protetores de preservação da musculatura esquelética durante o jejum e o que
vai acontecer é que a energia vai ser gerada principalmente pela degradação dos TG do
tecido adiposo através dos ácidos graxos que são liberados pelo tecido adiposo, enquanto
que os aas provindos dos músculos vão ainda continuar mantendo uma taxa de
neogliconênese basal para que possa ser suprida a demanda de glicose pelos tecidos que
utilizam glicose preferencialmente (eritrócitos, medula renal e SNC - retina).
Então a lipólise
continua durante o jejum liberando ácidos graxos que serão transportados pela albumina
até os tecidos e esses ácidos graxos são captados na sua maioria pelo próprio fígado, onde
eles são convertidos em acetil-coa e este acetil-coa normalmente entraria no CK, mas nós
estamos numa situação metabólica onde os intermediários do ciclo de Krebs estão sendo
captados para via de Gliconeogênese, portanto, a concentração dos intermediários e
principalmente de oxaloacetato é mais baixa, e o acetil-coa não vai para o CK, começa a se
acumular e acaba seguindo uma via do metabolismo secundário para formação de Corpos
cetônicos (aceto-acetato, beta-hidroxi-butirato e acetona).

Esses corpos cetônicos são
então exportados para o sangue e no sangue vão circular e serão captados por tecidos
como coração, músculo esquelético, rim e até mesmo cérebro. Nesses tecidos os corpos
cetônicos captados são biotransformados em acetil-coa e essa acetil-coa é que entra no
ciclo de krebs. Isso é possível porque nestes tecidos extra-hepáticos não ocorre a
gliconeogênese, portanto, o CK está ativo e o acetil-coa pode ser utilizado para gerar
energia na célula.
Gráfico: no ínicio a glicose provém de fonte externa, ou seja, exógena (é a glicose que está
sendo absorvida).Uma vez esgotado esse suprimento, a glicogenólise hepática mantém os
níveis de glicose sanguínea e ao mesmo tempo já inicia a gliconeogênese (lembrando que a
síntese de glicose pela gliconeogênese utiliza como precursor os esqueletos carbônicos dos
aas resultantes da proteólise muscular como já falado na fase de absorção). Então na fase
3 é a gliconeogênese que é a principal fonte de glicose sanguínea). Após extinguir toda
reserva de glicogênio hepático a gliconeogênese é a única fonte de glicose. então no jejum
e jejum prolongado é a gliconeogênese que mantém glicemia.

Gráfico: Esse gráfico evidencia que a gordura corporal é a nossa principal defesa
contra a privação alimentar. O que a gente observa é que a reserva energética armazenada
de glicogênio acaba rapidamente, então glicogênio mantido como reserva energética
corporal dura cerca de um dia aproximadamente. As outras reservas são utilizadas logo no
início da privação. Tanto as proteínas quanto as gorduras são utilizadas logo nas primeiras
horas de jejum, mas chega um momento que as proteínas começam a ser poupadas porque
como a gente já falou caso o animal continue usando os aminoácidos para sintetizar glicose
livremente ele vai sofrer uma grande depressão de proteínas musculares e isso vai trazer
problemas até mesmo locomotores. Então o que acontece é que o organismo começa a
privar o consumo de proteínas e priorizar o consumo de ácidos graxos e lipídeos. É claro
que isso é às custas da produção de Corpos cetónicos e que muitas vezes podem causar
transtornos no sistema nervoso uma vez que os corpos cetónicos são neurotóxicos. Por
último, realmente o organismo começa a abrir mão da preservação das proteínas e começa
a utilizar elas para degradação (6-7 semanas de jejum - fisiologicamente não acontece).
Referências: Aula ministrada dia 17/03 pela Professora Adriana Pedrenho
Fonte das imagens: Manual de Bioquímica com correlações clínicas. Devlin, 2011
Cunninghan - Tratado de Fisiologia Veterinária. Klein, B.G., 5°ed., 2014.

















Correo electrónico del Dr. Itua: drjekawo@gmail.com www.drjekawo.com y número de WhatsApp +2347059818667. Las medicinas herbales del Dr. Jekawo curaron completamente mi herpes y artritis, y han pasado 3 años sin ningún brote. Estoy muy feliz de estar finalmente curado.
ResponderExcluirEl Dr. Jekawo puede curar las siguientes enfermedades: Herpes, VIH, EPOC, Diabetes, Parkinson, Hepatitis, Cáncer, Enfermedad de Lyme.